KEYWORDS
Antibiotics, diabetes, gut microbiota, inflammatory bowel disease, internal medicine
INTRODUCTION
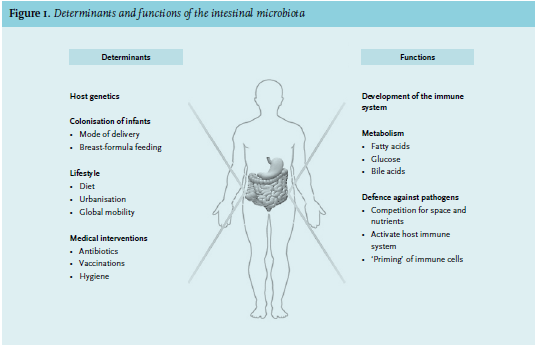
The field of microbiota research has exploded in recent years. In the first six months of 2014, over 100 peer-reviewed articles were published on this topic on each consecutive day. The awareness that commensal microorganisms are not simple bystanders in our bodies, but instead play key roles in physiology and pathology, has excited scientists and clinicians in almost every discipline of medicine.1-3 The unprecedented success of faecal transplantation as a potential cure for recurrent Clostridium difficile infection has become the gem of the microbiota research field;4 still, the gut microbiota is suspected of having an important role in a whole range of diseases.5 ‘The last human organ’ plays a major role in the development of the immune system, the defence against pathogens and the metabolism of fatty acids, glucose and bile acids (figure 1).6 Of special interest is the degradation of otherwise non-fermentable dietary fibre such as resistant starch into short-chain fatty acids (SCFA), mainly by bacteria from the Bacteroidetes phylum.7 These SCFA – particularly acetic, propionic and butyric acid – have anti-inflammatory and immune-signalling properties and are an energy source for epithelial cells.8 Our gut contains ten times more bacteria than our bodies contain cells of our own.9 Moreover, the collective microbiome is 150 times larger than the human genome, indicating the astonishing number of processes that the intestinal microbes are involved in.9 It remains to be determined, however, if this new wealth of knowledge will affect clinical medical practice. In this review we provide an overview of the current literature on the role of the gut microbiota in health and disease from the perspective of the internist.
COMPOSITION OF THE GUT MICROBIOTA
Determining the exact composition of the gut microbiota has been difficult due to the limited success of culturing (often anaerobic) bacteria. With the development of specialised microarrays and high-throughput sequencing techniques, our knowledge on this subject has started to expand quickly (textbox).9,10 The new insights gathered with these new techniques start right at birth: we now know that we are not born sterile, as low levels of bacteria are detected in meconium and umbilical cord blood.11 Major colonisation, however, starts after birth and is influenced by many factors such as the genetics of the host, mode of delivery, breast or formula feeding, nutrition and antibiotics. The diversity of the gut microbiota increases in the first years of life,11 after which the core composition appears to remain relatively constant during adult life:12,13 60% of all gut microbiota strains in healthy adults remain detectable over a period of five years, following a power law that suggests that this core is present for a much longer period.13,14 Each individual has their own unique microbiota. Even between healthy persons the composition may vary strongly as was shown in over 1000 adults.10,15 However, when looking at a functional level, most people carry equal amounts of bacterial genes involved in metabolic pathways. This suggests that the microbiota as a functional organ is similar among human beings.10 Diet is one of the most important determinants of the microbial composition.16 A study in healthy adults showed that an extremely fat-rich diet is capable of changing the microbiota in just a few days.17 Still, pinpointing causal relationships between the absence or overabundance of bacterial strains and clinical observations is difficult. Few prospective and intervention studies have been performed hitherto.13,17,18 With older age, inter-individual differences in gut microbiota composition become larger. For instance, long-term care residency is associated with lower bacterial diversity.19 Still, the largest and most direct effects on microbial composition are achieved by antibiotics.
INFLUENCE OF ANTIBIOTICS
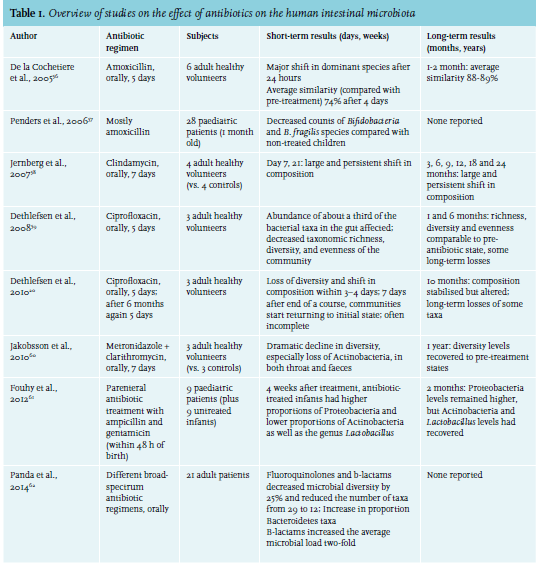
A simple antibiotic course, prescribed to kill just one or a few pathogens, will quickly result in major disturbances of the gut microbiota (table 1).20 Recovery and stabilisation of its diversity level may take months, and the new composition of bacteria may significantly differ from pre-treatment. Not only bacteria that fall within the spectrum of the antibiotic are affected; species that are dependent on the ones being killed may disappear as well. Vancomycin, for example, does not only lead to the disappearance of Gram-positive species, but also Gram-negative species and results in more dramatic shifts than amoxicillin.21,22 Antibiotics may thus have negative effects on the health of patients, for example on their metabolic state. Use of antibiotics has been associated with the development of obesity, diabetes mellitus and asthma.2
THE ROLE OF THE GUT MICROBIOTA IN DISEASE
Gastrointestinal infections
A healthy gut microbiota protects the host directly against pathogens such as C. difficile, by competition for nutrients, space and binding spots on the epithelium and production of bacteriocins, but also indirectly, by activation of the host immune system resulting in release of IgA, cytokines and antimicrobial peptides.23 Depletion of the microbiota by antibiotics therefore creates an opportunity for C. difficile to proliferate. Multi-drug resistant organisms are a rapidly increasing problem around the world. Vancomycin-resistant enterococci (VRE) are now a leading cause of bloodstream infections in haematopoietic stem cell transplant recipients.24 In both mice and men, domination by VRE (being at least 30% of the microbiota) correlates with subsequent development of a bloodstream infection25 and in mice can be reversed by faecal microbiota transplantation (FMT).24 This suggests that faecal transplantation therapy might be effective in both prevention and treatment of VRE infections. Assessment of faecal microbiota may be used to identify those patients at highest risk for bloodstream infection.25 The first randomised clinical trial for FMT in recurrent C. difficile infection was stopped prematurely because of a clear advantage in the FMT arm: 15 of 16 patients had complete resolution of disease, compared with four of 13 patients in the vancomycin arm and three of 13 patients in the vancomycin + bowel lavage arm.4 European recommendations now include FMT for patients with multiple relapses of C. difficile infection.26

Inflammatory bowel disease
Inflammatory bowel disease (IBD) is hypothesised to be either an aberrant immune response against normal commensal bacteria, or a normal immune response against abnormal gut microbiota, both driven by an autoimmune genetic background.27 Indeed, IBD patients have a significantly lower microbial diversity compared with healthy controls28 as well as a lower abundance of immune-modulating bacteria such as Faecalibacterium prausnitzii and Akkermansia muciniphila.5,29 F. prausnitzii and A. muciniphila produce butyrate and propionate, respectively, both SCFA that are thought to protect the mucosal barrier. A meta-analysis of 41 cases in which FMT was used for ulcerative colitis found that 63% of patients entered remission and 76% could stop medications.28 Obviously, these numbers may suffer from publication bias. Thirteen clinical trials are currently ongoing to evaluate the effect of FMT in IBD (table 2).

Transplant immunology
Intestinal inflammation secondary to graft-versus-host disease (GVHD) is likely to be influenced by the gut microbiota. A mouse model of GVHD caused by allogeneic stem cell transplantation (SCT) showed that mice developing intestinal GVHD have a decreased bacterial diversity compared with those that do not.30 A greater proportion of bacteria consisted of Lactobacillales, while Clostridiales were decreased. The same pattern was found in 31 patients receiving allogeneic SCT, probably due to chemotherapy, irradiation and antibiotics. The percentage of Enterococcus faecium and Enterococcus faecalis (of the order Lactobacillales) in the microbiota was 21% in patients who did not develop intestinal GVHD, 46% in those that subsequently developed GVHD and 74% at the time of actual GVHD.31 The diversity level at the time of engraftment seems to predict outcome: after classifying 80 SCT patients into low, intermediate, and high diversity groups, overall three-year survival was 36%, 60%, and 67% respectively.32
Colorectal cancer and response to chemotherapy
The intestinal microbiota play a key role in mucosal immune responses, epithelial cell homeostasis, barrier function and metabolism, all of which are important for tumour formation. For example, the microbiota produce (precursors of) carcinogenic substances, such as nitrate.5 A strictly animal-based diet (meats, eggs and cheeses) of only five days in healthy volunteers increased bile acids produced by the microbiota, including deoxycholic acid which promotes DNA damage and hepatic carcinoma.17 Accordingly, bile-tolerant bacteria such as Bilophila wadsworthia were increased, which in mice have been described to induce IBD by producing hydrogen sulphide.17 This supports the association between diets that are rich in red meat and the development of colorectal cancer.33 Faecal transplantation from colon tumour-developing mice to germ-free mice resulted in a higher incidence of colon tumours in the latter, compared with transplantation from control mice – most likely by inducing inflammation prior to induction of tumorigenesis by dextran sodium sulphate in drinking water.34 The gut microbiota also influences the effectiveness of anticancer therapy.35-37 In different cancer mouse models, it was shown that in the absence of a healthy microbiota, tumour-infiltrating myeloid cells are less capable of producing cytokines and inducing cytotoxicity upon immunochemotherapy or platinum chemotherapy, resulting in decreased necrosis of tumours.35 Both cytotoxic drugs and irradiation may cause translocation of bacteria or bacterial particles to lymph nodes, thereby enhancing immune responses directed against malignant cells.36,37 Destruction of the microbiota by antibiotics and other drugs may therefore reduce the efficacy of these treatments.35-37
Allergic and infectious airway diseases
Low diversity of the gut microbiota, whether caused by low exposure to environmental microbes or (prenatal or postnatal) use of antibiotics, is associated with increased risk of allergic airway disease.38,39 The intestinal microbiota is postulated to enhance the haematopoiesis of dendritic cell precursors in bone marrow by production of SCFAs.40 Intriguingly, dendritic cells from mice that have received propionate exhibit an impaired ability to activate T-helper 2 cells in the lung, thus preventing allergic airway inflammation.40 Supplementation of dietary fibre or SCFA may therefore prevent or reverse allergic airway disease. A similar ‘priming’ effect by healthy gut microbiota has been observed in murine models of pneumonia and peritonitis.41-43 Depletion of the gut microbiota by antibiotics in neonatal mice appears to hamper neutrophil production by the bone marrow, thus leading to increased counts of pathogenic bacteria and decreased survival.43 It is suggested that lipopolysaccharide from gut bacteria is the priming component, although another role for SCFA is not excluded. For now, evidence for this ‘gut-lung axis’ in humans is lacking.
Metabolic disorders
Obesity has been associated with a lower microbial diversity in the intestine.44,45 Studies have reported an association between obesity and increased levels of Firmicutes, combined with decreased Bacteroidetes (the two most dominant phyla), although this could not be replicated in all cohorts.5,46 Overweight and obese individuals have higher levels of SCFA, without a different intake of dietary fibre, reflecting either an increased production or decreased absorption by the gut and utilisation by bacteria.46 In both mice and men, there is evidence that the obese microbiota is more effective at harvesting energy from food, as suggested by enriched biochemical pathways.5,47 Accordingly, transplantation of gut microbiota from obese to germ-free mice leads to more weight gain than transplantation of microbiota from control mice.47 Transplantation of faeces from lean human donors into patients with metabolic syndrome decreases peripheral insulin resistance six weeks after transplantation. This is potentially caused by increased levels of butyrateproducing bacteria (Roseburia intestinalis).48 By contrast, among 20 children of 3-5 years of age, levels of butyrateproducing bacteria were also found to be higher in those that subsequently developed type 1 diabetes.49 Of interest, seven days of vancomycin therapy resulted in decreased peripheral insulin sensitivity in ten males with metabolic syndrome.22 This was associated with a decrease in Firmicutes bacteria and decreased metabolism of bile acids.
From theory to therapy
Although the role for the gut microbiota in the pathophysiology of disease may be easy to imagine, translation of this knowledge into new therapeutic strategies represents a tremendous challenge. Changing one’s intake of specific nutrients would be the most natural way to prevent diseases. Consumption of dietary fibre seems one of the best solutions, since they are fermented into SCFA that may have beneficial effects. Both extreme diets17 and the use of various fibres were shown to modify the human intestinal microbiota, although the latter not in all subjects.50 Probiotics that include a variety of lactic acid bacteria are by now commonplace in both the supermarket and the clinic, but their effectiveness is highly debatable.51 Moreover, higher mortality in patients treated with lactic acid bacteria during severe pancreatitis compared with placebo treatment has led to increased caution regarding these bacteria, notably in a clinical setting.52 The successful trial on severe C . difficile infection has put the spotlight on FMT.28 However, we currently do not know how to select a microbiota that is best to transplant. In addition, the possibility of transplanting pathogenic microorganisms, or the possible risk of metabolic and autoimmune diseases, might be a reason for caution. So far, no serious (long-term) adverse events have been reported. Transplanting a cocktail of bacteria (the synthetic microbiota) that can be cultured or isolated easily, may be a key objective the coming years.53 The first steps were already taken in 1989, when ten selected intestinal strains that showed inhibitory effects against C. difficile cured five patients with C. difficile infection.53,54 More recently, two C. difficile infected patients were transplanted with 33 cultured bacterial species, isolated from healthy donor faeces – both resulting in resolution of disease and no relapse in 24 months.23 These results encourage the development of synthetic, standardised microbiota.
CONCLUSION
The booming popularity of the gut microbiota in biomedical research seems justified; in fact, the interest in host-microbiota interactions and their potential value as a therapeutic target seems rather belated in view of our ancient commensal relationship. Obviously, results from animal studies cannot be translated directly to human health and disease. Still, some of the possible microbiotadisease interactions will prove to be relevant and applicable in everyday clinical practice. The great possibilities offered by this new field of research provide hope for new, relatively straightforward and inexpensive therapies. However, most research is currently in a preclinical phase, and causative relationships remain to be established. Moving from correlation to causality will be particularly difficult for phenotypically and/or genetically heterogeneous disorders, such as diabetes, IBD and GVHD.55 In anticipation of gut microbiota-targeted therapies, it seems important to realise the key function of the gut microbiota in physiological processes and the potential collateral damage we cause when disrupting this well-balanced ecosystem.
DISCLOSURES
The authors declare no conflicts of interest. Financial support: W.J. Wiersinga received a Clinical Fellowship from the Netherlands Organisation for Health Research and Development (ZonMw).
REFERENCES