KEY WORDS
Microbiota, hematologic malignancies, antibiotics, immune activation
INTRODUCTION
Overview of the human microbiota: characterization and regulation of microbiota
Human beings are inhabited by innumerable microorganisms that interrelate with the host in a reciprocal way, establishing a combined and efficient ecosystem – the microbiota- that can affect healthiness as well as disease.1,2
The term ‘gut microbiota’ indicates the masses of microorganisms living in the intestine. Most microorganisms inhabit the more distal portions of the intestinal area, where their bio-mass exceeds 1011 cells per gram content.3 Nevertheless, all cavities that connect to the outside and body surfaces are populated by mutable and personalized ecosystems of viruses, fungi, archaea, bacteria and protozoa.
The relationship between the host and the microbiota is symbiotic. The host offers a vital habitat for the microbiome, whereas microorganisms participate in host health via synthesis of essential amino acids, short chain fatty acids (SCFAs) and vitamins.4-6
In a recent study, Sonowal et al. showed that small molecules related to indole and originating from commensal microbiota act in different phyla to augment healthy aging. The action of indoles on health span depends upon the aryl hydrocarbon receptor, a conserved sensor of xenobiotic small molecules. In older animals, indole stimulates genes associated with oogenesis and, accordingly, prolongs reproductive span. These results are improving efforts of developing therapeutics based on microbiota-derived indole to decrease frailty in humans.7
The microbiota is distinguished by its unremitting dynamic renewal of microbiota.8 The different microbiota inhabiting human surfaces are not casually developed. They are the result of several elements, such as age, environmental conditions, lifestyle, smoking habit, antibiotics therapy, genetic factors and contact with pathogenic organisms.9-20 A substantial change in diet modifies microbial configuration within just 24 h of commencement, with a return to baseline two days after diet suspension.21 Moreover, the gut microbiome of animals nourished with a high-sugar or high-fat diet is more disposed to circadian rhythm disruption.22
Numerous diets, including vegan, gluten-free, omnivore, Western and Mediterranean, have been investigated for their ability to regulate the gut microbiota. In numerous studies, a diet high in animal fat and protein, but low in fiber, causes a pronounced reduction in numbers of beneficial Eubacterium species and Bifidobacterium.23-25 Furthermore, host circadian clock and hormonal status alter gut microbial ecology through nourishment and diurnal rhythms; jetlag and long-distance voyage cause the disturbance of this clock, and can therefore alter the gut microbiota.26,27
The initiation of novel tools has strongly impacted the interpretation of the controlling systems by which microorganisms and hosts interrelate to provoke a health or disease condition in the host. Next generation sequencing and methods connected to metabolome analysis, such as mass spectrometry, are critical for evaluating the microbiota structure and investigating the metabolic, functional and genetic action of the microbiota.28-30
Carcinogenesis and microbiota
Oncomicrobes comprise organisms that can directly injure DNA and modify host cellular processes.31-33 Several well-recognized oncomicrobes are viruses, which introduce oncogenes into host genetic material. Interestingly, several bacteria have elaborate competitive tactics which can harm DNA of contending organisms. Unfortunately, these same processes can also modify host DNA, resulting in mutations and perhaps, to carcinogenesis. Bacterial DNA can incorporate into human genomes, principally the mitochondrial genome, through an RNA intermediate, and this occurs more commonly in cancerous rather than healthy tissues.34 DNA mutations may also be caused by toxins generated by bacteria,35,36 and bacterial proteins can initiate signaling actions in host pathways that control cell growth.37,38 Nevertheless, few microorganisms are identified as oncomicrobes (see table 1). This may partly be due to difficulties in recognizing microorganisms as the causal mediators of carcinogenesis. The causal agent may be absent from the cancer site due to an environmentallydriven population of organisms, or the microbe may have started host cellular injury by a “hit and run” action after only short interaction with the host tissue.
Nevertheless, few microorganisms are identified as oncomicrobes (see table 1). This may partly be due to difficulties in recognizing microorganisms as the causal mediators of carcinogenesis. The causal agent may be absent from the cancer site due to an environmentallydriven population of organisms, or the microbe may have started host cellular injury by a “hit and run” action after only short interaction with the host tissue.
There is increasing evidence that the conformation of the microbiome may influence, and is controlled by, the human immune system.39 An unbalanced microbial structure been connected with reduced immune competence, predisposition to infections and inflammatory diseases.40,41 Experiments performed using germ-free animals propose that microbiota directly stimulate local intestinal immunity through their actions on toll-like receptor (TLR) expression,42 differentiated T cells, antigen presenting cells and lymphoid follicles,43,44 as well as by modifying systemic immunity by augmented systemic antibody production and splenic CD4+ T-cells.45,46
Microorganisms and microbial elements such as lipopolysaccharide (LPS) can up-regulate TLRs, which can provoke an activation of nuclear factor-kB (NF-kB), which is critical for controlling tumor-associated inflammation,47,48 invasion, growth, survival and immunosuppression.49 Bacterial LPS has also been demonstrated to hasten cell proliferation by c-Jun N-terminal Kinase activation.50
Remarkably, T-helper cell 17 (Th17) differentiation from naïve T-cells seems to be dependent on the segmented filamentous bacteria. Experiments have demonstrated that Th17 are lacking in the small-intestinal lamina propria of germ-free mice, which is their primary differentiation location, while modifications in the gut microbiota are closely related to important variations in Th17/Regulatory T-cell (Treg) balance, possibly mediated by epigenetic mechanisms. This is proven by emergent data connecting an unbalanced microbial structure to epigenetic modifications.51-53
Administration with segmented filamentous bacteria provoked an augmentation in production of interferon (IFN) gamma, interleukin-10 (IL-10), and IL-17,54 while administration of Sphingomonas yanoikuyae provoked a modification in immune cells. Bacterioides fragilis induces a Th17 response in animals, which was then demonstrated to be necessary for tumorigenesis, while administration with human commensal bacterium Bifidobacteriom longum, Bacteroides thetaiotaomicron or both caused an increase in the production of the IFN-gamma- and TNF-alpha pathways.55 As previously mentioned, gut flora can also control host immunity by epigenetic changes. For example, microbialoriginated butyrate reduces histone deacetylases 6 and 9, which results in higher numbers of Treg cells and enhanced acetylation in the promoter of the FOXP3 gene.56,57 Metabolic relationships and dietary elements can in some way induce tumor expansion. Metabolic end products may include pro-carcinogenic factors that promote cancer growth.58 Contact with these elements, referred to as the “exposome”, can influence oxidative stress and DNA stability in a host, increasing the danger of developing tumor.59,60 (see figure 1). Several carcinogenic factors can be produced by the cometabolism of xenobiotics by bacterial β-glucuronidases.61 Significant examples comprise the metabolism of azoxymethane62 and irinotecan.63 Similarly, production of harmful metabolites is connected with microbial catabolism of dietary proteins. These putrefactive activities in the bowel provoke the production of N-nitroso elements that cause DNA damage via alkylation.64,65 The metabolism of aromatic amino acids also causes the production of phenols, indoles, p-cresol and phenylacetic acid.66 Polyamines are a diverse group of toxic elements and catabolism of the main polyamines is connected with oxidative stress and tumors67 (see table 2).
As previously mentioned, gut flora can also control host immunity by epigenetic changes. For example, microbialoriginated butyrate reduces histone deacetylases 6 and 9, which results in higher numbers of Treg cells and enhanced acetylation in the promoter of the FOXP3 gene.56,57 Metabolic relationships and dietary elements can in some way induce tumor expansion. Metabolic end products may include pro-carcinogenic factors that promote cancer growth.58 Contact with these elements, referred to as the “exposome”, can influence oxidative stress and DNA stability in a host, increasing the danger of developing tumor.59,60 (see figure 1). Several carcinogenic factors can be produced by the cometabolism of xenobiotics by bacterial β-glucuronidases.61 Significant examples comprise the metabolism of azoxymethane62 and irinotecan.63 Similarly, production of harmful metabolites is connected with microbial catabolism of dietary proteins. These putrefactive activities in the bowel provoke the production of N-nitroso elements that cause DNA damage via alkylation.64,65 The metabolism of aromatic amino acids also causes the production of phenols, indoles, p-cresol and phenylacetic acid.66 Polyamines are a diverse group of toxic elements and catabolism of the main polyamines is connected with oxidative stress and tumors67 (see table 2). 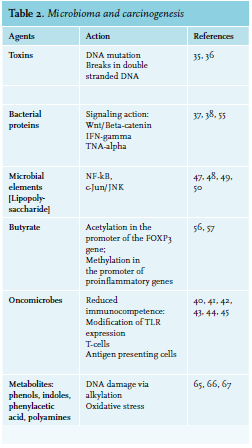
Chemotherapy and microbiome
Chemotherapy can harm normal cells of the intestinal system and possibly cause gastro-Intestinal (GI) problems.68 The cytotoxic actions of these therapies provoke a supplementary immunosuppression, which causes febrile neutropenia and bloodstream infections. Furthermore, the usage of prophylactic and therapeutic antibiotics alters the GI microbiome.69
Galloway-Peña et al. studied intra-patient temporal microbiota changeability and its clinical impact on tumor subjects during chemotherapy.70 These patients presented an elevated level of intra-subject temporal instability of oral and fecal bacterial multiplicity. Days on antibiotics were substantially connected with extended temporal variability of oral bacterial multiplicity and microbiome structure. These results indicate that an increased variability was connected with adverse clinical prognosis. Moreover, the data show the relevance of longitudinal microbiome analyses.
Unfortunately, direct interaction with bacteria can influence the effectiveness of chemotherapeutic agents.71 Mass spectrometry and high-performance liquid chromatography analysis demonstrated that contact with the bacteria determined a biotransformation of certain chemotherapy drugs. The ability of bacteria to reduce the antitumor effect of gemcitabine and to increase that of the prodrug CB1954 was demonstrated in vivo.72 Hence, data in experimental models propose a complicated interaction between numerous chemotherapeutic agents and microbiota.
Exhaustive mechanistic studies in vivo have been reported for platinum compounds and cyclophosphamide (CTX).73,74 CTX, prescribed for therapy of hematologic diseases and solid cancers, harms the small intestine epithelium causing migration of several microorganisms into lymph organs. This barrier breech provokes a T-helper cell-mediated anticancer response and augments drug effectiveness.73 The antitumor action of platin treatment is radically reduced in germ-free mice or in mice in which gut bacteria have been reduced by broad-spectrum antibiotics.74 Combined with experimental results that Lactobacillus acidophilus supports the anticancer action of cisplatin, clinical data propose the possibility that probiotic bacterial species may promote antitumor action while inhibiting some of the toxic side effects of particular drugs75-77 (see table 3). 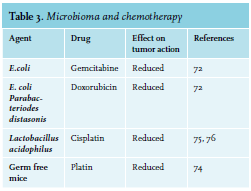 The intestinal chemotoxicity of methotrexate is due in part, by activation of TLR4 by microbial products such as Cif, as secreted toxin from Pseudomonas aeruginosa. Activation of TLR2 protects the mucosa against methotrexate-induced damage by augmenting the expression of the ABC transporter multidrug resistance protein 1 (MDR1; also known as P -glycoprotein and ABCB1), which controls the efflux of xenobiotics from intestinal epithelial cells.78
The intestinal chemotoxicity of methotrexate is due in part, by activation of TLR4 by microbial products such as Cif, as secreted toxin from Pseudomonas aeruginosa. Activation of TLR2 protects the mucosa against methotrexate-induced damage by augmenting the expression of the ABC transporter multidrug resistance protein 1 (MDR1; also known as P -glycoprotein and ABCB1), which controls the efflux of xenobiotics from intestinal epithelial cells.78
Microbiota and immunotherapy
Improvements in treatments and immunotherapies have generally discounted microorganisms as a part of the tumor therapy, until recently. The intestinal microbiome has been discovered to modify responses to tumor immunotherapy.79
Vetizou et al.80 and Sivan et al.81 demonstrated that the efficiency of immune checkpoint inhibitors (ICIs) in therapy is reliant on the host microbiota and that the ICIs responded poorly in animals raised under germ-free situations. The authors discovered that, in the presence of microorganisms, host antigen-presenting cells stimulate IFN γ-generating T-cells. Vetizou et al. studied how the curative efficiency of cytotoxic T-lymphocyte-associated protein 4 blockade, as mediated by ipilimumab, might also be caused by elements of the gut microbiota.82
It is now clear that primary resistance to immune checkpoints can be attributed to an abnormal gut microbiome composition. Fecal microbiota transplantation (FMT) from cancer patients who responded to immune checkpoints into germ-free or antibiotic-treated mice ameliorated the antitumor effects of programmed cell death 1 protein (PD-1) blockade, whereas FMT from non-responding patients failed to do so. Oral supplementation with Akkermansia. muciniphila after FMT with non-responder feces restored the efficacy of PD-1 blockade in an IL-12-dependent manner by increasing the recruitment of CCR9+CXCR3+CD4+ T lymphocytes into mouse tumor beds.83 Moreover, in melanoma patients undergoing anti PD-1 immunotherapy, significant differences were observed in the diversity and composition of the patient gut microbiome of responders versus non-responders. Analysis of patient fecal microbiome samples showed significantly higher alpha diversity and relative abundance of bacteria in the Ruminococcaceae family in responding patients.84
Microbiota and bone marrow transplantation
Hematopoietic stem cell transplantation (HSCT) is frequently used as curative therapy in patients with hematological diseases. The microbiome could be implicated in local and systemic complications after HSCT. Antibiotic-caused loss of lower gut diversity has been suggested as an independent predictor of unfavorable outcome, while progress of graft-versus-host disease (GVHD) appears to be a main contributor to mortality.85,86 Lately, microorganisms of the genus Blautia were identified as the connection between modifications in microbiota structure, onset of GVHD and a favorable outcome.87,88 In fact, increased amounts of bacteria belonging to the genus Blautia were associated with reduced GVHD lethality. Blautia abundance was also associated with improved overall survival.87 The role of the intestinal microbiota and its potential influence on clinical outcomes for patients undergoing allogeneic HCT (allo-HCT) has been investigated in recent years.89-91
Acute GVHD (aGVHD) might reduce the chances of successful allogeneic bone marrow transplantation. Present pathophysiologic hypotheses of aGVHD include proinflammatory cytokines and bacterial LPS as main triggers for aGVHD. LPS originates principally from Gram-negative bacteria and can pass in circulation via the damaged mucosal barrier after the conditioning regimen. Probiotic microorganisms have been proven to modify the components of the intestinal microflora and thus mediate anti-inflammatory actions. It has been suggested that changing the enteric flora using the probiotic microorganism Lactobacillus rhamnosus GG would improve aGVHD.92,93 Analyses of fecal specimens taken from recipients of allo-HCT around the time of engraftment have shown that a reduced intestinal microbiome diversity is associated with significantly worse survival outcomes, and that the alterations in the microbiota composition may influence important clinical outcomes such as aGVHD and disease relapse.94-96
It has been hypothesized that approaches to restore a patient’s microbiome diversity after HCT may improve outcomes after HCT. Although approaches in restoring microbiome diversity are still investigational, FMT from a healthy individual promise to be a remarkably effective therapy.97 FMT refers to the infusion of fecal suspension from a healthy donor into the gastrointestinal tract of a patient in order to restore the microbiota and cure the disease. Manipulation of the intestinal microbiota by FMT may influence the immune system and improve immune-mediated enteritis such as gut aGVHD.98
Moreover, bloodstream infection (BSI) is one of the biggest causes of death among cancer patients. A study stated that gut domination, described as the occupation of at least 30% by a specific bacterial taxon, is connected with BSI in subjects undergoing allo-HSCT. These data suggest that the intestinal microbiota can recognize high-risk subjects before HSCT and that management of the intestinal microbiota to prevent BSI in high-risk subjects could be a useful treatment.99
Finally, a common side effect of myeloablative therapy used during HSCT treatment is GI mucositis.100 A model, created by Sonis, illustrated a process for bacterial infection due to GI mucositis after HSCT.101 It comprises an ulcerative stage with increased permeability and harm to the gut mucosal barrier. This stimulates microbial translocation, described as the migration ofmicroorganisms from the GI tract to extra-intestinal places, such as the blood.102
Antibiotics, microbiota and cancer
The discovery that a persistent usage of antibiotics can stimulate some tumors in cancers such as esophageal, gastric and pancreatic cancers,103 suggests an association between antibiotics, microbiota and cancer.
In fact, there is an indirect indication that an unbalanced microbial structure provoked by antibiotic treatment can augment the incidence of some tumors. An epidemiological study (125,441 cases and 490,510 matched controls) suggests that the use of antibiotics may also influence the occurrence of breast cancer (macrolides, penicillin, cephalosporins, sulphonamides and tetracyclines).103,104 These data propose that some antibiotics are carcinogenic (tetracyclines inhibit replication of mitochondrial DNA) or that they cause changes in the structure of the microbiota that promote the growth of cancers. However, these results have to be cautiously interpreted because they referred to solid tumors, and the iterative use of antibiotics may denote the existence of immune alterations that could be the main cause of microorganism infections and increased tumor frequency.
Nevertheless, antibiotics can enhance the growth of tumors in animals. This has been demonstrated for the therapy of proto-neu transgenic animals in which tumors appear with ciprofloxacin plus metronidazole.105,106
Diet, probiotics and microbiota
There is outstanding interest in the manipulation of the microbiome as a possibility to reduce the incidence of cancer. The intrinsic relationship between microbiome, diet and health demonstrates a possible improvement of our health if we modify our diet.107,108 Indeed, the American Cancer Society has indicated that diets might be responsible for 30% of tumor cases in developed states and 20% in developing nations.109
Reports indicate that diet modifies the structure of the gut flora, and this can impact on the immune response.110,111 On the other hand, eating particular types of foods (fish, fruits, poultry and vegetables) may prevent several tumors.112-115
Metabolism of food can also cause the production of bioactive molecules with chemo-protective activities. Carbohydrate fermentation provokes the production of the short chain fatty acids acetate, butyrate and propionate. These substances can act with free fatty acid receptors in the gut epithelia to influence immune processes, such as the production of cytokines.116 Other researchers have demonstrated that butyrate can increase the number of formations of tight junction proteins through the modulation of AMP-activated protein kinase.117 Butyrate also has anti-carcinogenesis actions, principally by two means: the stimulation of G-protein-coupled receptors 41 and 43 and the reduction of histone deacetylase. Some of the described actions of butyrate are improvements of specific pro-apoptotic gene expression in tumor cells and the reduction of the pro-inflammatory pathway of NF-kB.118,119
Moreover, there is epidemiological evidence supporting chemo-protection derived from the intake of vegetables and fruits, which have been ascribed to secondary metabolites, such as polyphenols and glucosinolates.120 Polyphenols are powerful anti-oxidants, but experiments have demonstrated that they are not able to achieve bioactive levels in the blood.121,122 These elements can also modulate immune responses by modification of the intestinal microbial structure.123,124
Probiotics can live in the gut and stimulate the recuperation of regular intestinal microbiota; the most well-known probiotics are Bifidobacteria and Lactobacillus. 125
Probiotics increase the immune response through augment of natural killer cell (NK) cytotoxicity and the stimulation of phagocytes.126 The effectiveness of NK cells can also be augmented when a mixture of probiotics and dextran is utilized.127 Furthermore, the increase of probiotics seems to increase the production of immunoglobulins IgM and IgA, thus fortifying the adaptive immune response.128-132
Bifidobacteria and Lactic acid bacteria are able to induce the secretion of elements that reduce inflammation by downregulating IL-8 production, NF-kB-dependent gene expression and concentrations of macrophage-attracting cytokines.133-136 Morita et al. also demonstrated a significant augmentation of the expression of IL-6, IL-10, and IL-12 after the stimulation of macrophages with Lactobacillus acidophilus.137 Nutritional supplements with probiotics have been used to increase immune system activity in elderly people.138,139 Li et al. showed that Prohep - a new probiotic mix - reduces cancer development in an animal model.140 Probiotics change the intestinal bacterial structure so that it comprises specific advantageous microbes, such that Oscillibacter and Prevotella, known fabricators of anti-inflammatory substances, which are able to decrease Th17 polarization and stimulate the differentiation of Treg/ Tr1 cells in the intestine.
Microbiota and hematologic malignancies
Acute Lymphoblastic Leukemia (ALL)
Rajagopala et al. compared the GI microbiota constitution of adolescent and pediatric leukemia subjects with their healthy relatives.141 They identified modifications in the microbiota composition of leukemia subjects during chemotherapy by evaluating samples taken before and after treatment at variable time points throughout the treatment. Their results supply relevant data on GI microbiota structure in immunocompromised patients and suggest that the baseline microbiota of these patients is significantly different from their healthy siblings.
The microbiota structure of patients and siblings are dominated by components of Prevotella, Bacteroides and Faecalibacterium. The microbiota diversity of the patient groups was substantially lower than that of the controls. It was possible to differentiate between the leukemia subjects and the controls based on their microbiota composition. The principal taxa comprise Roseburia, Ruminococcus2, Anaerostipes and Coprococcus with moderately higher abundance in the controls.141
Information regarding the oral microbiota in leukemia subjects is lacking and mostly inadequate. Among some patients, leukemia first presents in the oral cavity.142,143 Oral symptoms that commonly arise in leukemia subjects include gingival enlargement and bleeding, candidiasis, oral ulceration and periodontitis144-146 and oral microorganisms are implicated in the onset of such problems.147 Specific oral microorganisms have been shown to contribute to septicemia, which might delay antineoplastic therapy or even put the subjects’ life at risk.148-153 Consequently, a suitable therapy for oral lesions could result in a more satisfactory outcome of both oral and systemic complications.
However, Wang et al. studied the structure of the supragingival plaque microbiota of ALL pediatric subjects and of the healthy controls.154 The oral microbiota of leukemia subjects had less diversity related to controls. Microorganisms grouped into two main clusters, patients and controls, with diverse composition. Variations of specific taxa comprising the families Aerococcaceae and Carnobacteriaceae, Phylum Firmicutes, the order Lactobacillales, the genera Abiotrophia and Granulicatella and the class Bacilli were correlated with leukemia status. Nevertheless, it was demonstrated that the complexity of oral microbiota was not significantly diverse between leukemia subjects and controls until beginning of antineoplastic therapy. This contradiction might be ascribed to the methods employed to study oral microbiota. At the time of the study, oral microorganisms might have already undergone selection after ALL was detected, with some microorganisms inhibited and others prospering.155
However, the studies of oral microorganisms in ALL subjects, however, provided the possibility of recognizing potential microorganisms correlated with systemic infections in leukemia subjects. Results propose two taxonomical lineages (Firmicutes/Bacilli/Lactobacillales/Aerococcaceae/Abiotrophia, and Firmicutes/ Bacilli/Lactobacillales/Carnobacteriaceae/Granullicatella) that are much more copious in the supragingival plaque of ALL subjects than controls. This suggests that advantageous situations existed for their growth in the oral space of ALL subjects and could be responsible for an increased risk of bacteremia in leukemia subjects.
Abiotrophia and Granulicatella have been involved in endocarditis, otitis media, central nervous system infections, cholangitis and arthritis.156-159 Previous researchers have found that oral microorganisms are responsible for local infections and for 25-50% of systemic infections.160
Finally, adult survivors of ALL have health problems that arise years after termination of treatment. Chua et al. evaluated the anal microbiota structure of adult survivors of childhood ALL and controls. They recognized a modified population with decreased microbial diversity in tumor survivors, who also display signals of immune alteration comprising enhanced T-cell activation. The microorganism population among ALL survivors was enriched for Actinobacteria and depleted of Faecalibacterium, consistent with corresponding plasma levels of C-reactive protein and IL-6 and HLA-DR+CD4+ and HLA-DR+CD8+ T cells. They established a relationship between dysbiosis and immune alteration in adult ALL survivors. Actions that could reestablish microbial diversity may improve development of late effects of childhood ALL survivors, particularly chronic inflammation-related comorbidities.161
Acute myeloid leukemia (AML)
Bacterial infections and their complications are one of the most frequent and critical treatment-related toxicities in subjects with AML. Gram-positive cocci, principally the heterogeneous Viridans streptococci, are the most usually isolated microorganisms in patients with AML.162 However, many elements of the microbiota could play a positive role towards leukemic disease. The oral bacterium, Aggregatibacter actinomycetem comitans, generates a leukotoxin (LtxA) that is specific for white blood cells by interacting with lymphocyte function antigen-1 (LFA-1) on susceptible cells. Kachlany et al. valuated the in vitro and in vivo anti-leukemia action of the toxin. LtxA destroys malignant cell lines and primary leukemia cells from AML subjects, while healthy cells are moderately resistant to LtxA-mediated cytotoxicity. Levels of LFA-1 in Jurkat cell lines correlated with killing by LtxA and the toxin especially destroyed cells presenting the activated form of LFA-1. In a severe combined immune deficiency mouse model for human leukemia, LtxA had powerful therapeutic action resulting in long-term survival of the LtxA-treated mice.163
Interestingly for patients with erythroleukemia, a rare form of AML, kefir, a beverage obtained by the incubation of kefir grains with raw milk may be an effective therapy. Kefir grains are a symbiotic complex of diverse kinds of yeasts and bacteria, especially lactic acid bacteria, which congregate in a mostly carbohydrate matrix, called kefiran. In recent years, the action of kefir on some cancers has been investigated. Jalali et al. demonstrated that kefir caused apoptosis and necrosis in an acute erythroleukemia cell line (KG-1), by reducing growth. The study suggested that kefir may have the potential to be an effective therapy for erythroleukemia.164
Furthermore, numerous bacterial toxins are being investigated as potential anti-leukemia agents, either for their direct effects or to release therapeutic proteins against leukemia. LukS-PV, an element of Panton-Valentine leukocidin (PVL) produced by S. aureus, has certain anti-leukemia actions such as inducing leukemia cell differentiation and apoptosis, thus making LukS-PV an encouraging novel treatment strategy for leukemia.165 PVL is a staphylococcal synergohymenotropic exotoxin belonging to the pore-forming toxin family. PVL causes lysis of human polymorphonuclear neutrophils, monocytes and macrophages. Several works have proven that LukS-PV is able to cause leukemia cell differentiation.166 LukS-PV provoked differentiation by stimulating the extracellular-signal-reduced kinase (ERK) signaling pathway and c-JUN/c-FOS in human acute myeloid leukemia cells.167,168
B and T lymphomas
Adolescent/young adult Hodgkin lymphoma (AYAHL) is associated with reduced exposures to infections. Similarly, a study of AYAHL survivors suggested fewer early childhood fecal-oral exposures compared with health controls, and patients have immunological alterations. AYAHL is related to suppressed Th1 activity and an increase of Th2 response.169
Extension of gut microorganisms during childhood170,171 correspond with a change from a Th2 to a mature Th1-governed immune profile.170 Increased concentrations of Th2 cytokines and IgE in AYAHL subjects reduced cytotoxic T-cells and NK cells in Hodgkin lymphoma172 suggest the failure to make this Th2-to-Th1 change. These data suggest the possibility that the gut microorganisms may impact AYAHL.173,174 Cozen et al. explored whether fecal microbial diversity varied between AYAHL survivors and co-twin controls. In this small investigation, AYAHL survivors seem to have a reduction of rare gut microorganisms. Further study is required to clarify if decreased microbial diversity is a result of Hodgkin lymphoma, its therapy or a specific hygienic environment.173-176
About 12% of all human tumors are related to oncogenic viruses such as Epstein Barr Virus, Herpes Human Virus 8 and Human T-cell leukemia virus type 1.177 The occurrence of virus-related cancers, principally lymphomas, varies geographically and is induced by higher temperatures and environmental factors.178-201
Oxidative stress provoked by gut microorganisms can affect carcinogenesis and influence numerous pathways correlated with lymphomagenesis.195-201
Mucosal-associated lymphoid tissue (MALT) lymphomas are supposed to derive in the marginal zone and are connected with the presence of Helicobacter.202-205 This correlation was first revealed in an animal model infected with H. felis, an intimate relative to H. pylori and 154 days post-infection, 25% of mice had lymphoepithelial alterations while none of the controls did.206 An H. pylori infection was initially recognized in gerbils and displayed an augmentation in intestinal metaplasia.207 Since then, H. pylori infections have been recognized in animal models.208,209
H. pylori was classified as carcinogenic to humans (Group I carcinogen) in 1994 by an International Agency for Research on Cancer (IARC) Working Group based on the data from a small number of papers that studied gastric carcinoma.210 In 2009, a new Working Group evaluated significantly more results and confirmed that chronic infection with H. pylori is a Group 1 carcinogen with appropriate evidence that the infection causes gastric carcinoma and low-grade B-cell gastric MALT lymphoma.211
Gastric MALT lymphoma (GML) is strictly associated with H. pylori infection. In a retrospective evaluation of 144 consecutive patients admitted with GML, eradication treatment was extremely effective in causing complete remission (CR) and long-term prognosis was satisfactory. At multidisciplinary care stage EI, 92% of subjects received an H. pylori eradication therapy; 83% achieved CR after a mean period of seven months, and 86% remained in CR after a mean follow-up time of 105 months.212
A correlation was also hypothesized for other types of lymphomas. Numerous works described that most early-stage gastric diffuse large B-cell lymphoma (DLCBL) is H. pylori-dependent. Notably, DLCBL could possibly be treated by H. pylori eradication. Unlike MALT lymphoma, however, DLCBL may rapidly increase if it is unresponsive to H. pylori eradication. Consequently, detecting biomarkers that may predict an H. pylori-dependent status of gastric DLCBL is indispensable. Kuo et al. from Taiwan proposed that the expression of cytotoxin-associated gene A (Cag A) and CagA-signaling molecules p-SHP2 and p-ERK in malignant B cells is associated with H. pylori dependence.213 The same authors demonstrated that activating the B-cell-activating factor (BAFF) pathway upregulates NF-kB and causes BCL3 and BCL10 nuclear translocation in H. pylori-independent gastric DLCBL tumors with evidence of MALT. Moreover, they showed that the autocrine BAFF signal transduction pathway contributed to H. pylori independence in gastric MALT lymphomas without the t.11;18q21;q21 translocation.214
H. helmanii also contribute to MALT lymphoma which is preceded by endothelial venule-like vesicles, which are connected with lymphocyte enrollments.215 These models of microorganism-induced lymphoma however, appear to have variable results and may implicate bacterial and host elements.216,217
Other microorganisms such as Chlamydia psitacci, Campylobacter jejuni and Borrelia bergdorferi may also have an increase in the incidence of the disease in lymphoma progress.218
Infection of Borrelia burgdorferi may be causally related to B-cell non-Hodgkin lymphoma, as reported in one of two reports in Scandinavia.219,220Chlamydia psittaci, the agent of the zoonotic infectious disease psittacosis, has been found in MALT lymphomas in several non-gastrointestinal structures,221 while Streptococcus bovis has been connected with hematopoietic diseases such as chronic myelogenous leukemia, and chronic lymphocytic leukemia.222
Intriguing results originated from animals defective in the Ataxia telangiectasia-mutated gene (Atm-/- mice), which exhibit a high occurrence of thymic lymphoma.223 They are hypersensitive to modifications in microorganism content.224 Barlow et al. discovered that Atm-/- animals that were moved into more sterile situations lived longer and have a reduced lymphoma penetrance. In contrast, when they were relocated to standard specific-pathogenfree situations, their life and tumor latency decreased.225
Chronic Lymphocyte Leukemia (CLL)
A recent study in CLL subjects connected the efficiency of antineoplastic therapy with the use of antibiotics that alter intestinal microbiota. Pflug et al. assessed the effect of antibiotics on progression-free survival (PFS) and overall survival (OS).226 Among 800 CLL subjects, those receiving anti-Gram-positive antibiotics attained a substantially lower overall response rate (ORR). In the same study, authors evaluated patients with relapsed lymphoma. Of 122 patients with relapsed lymphoma, those treated with anti-Gram-positive antibiotics achieved a significantly lower ORR. Patients with anti-Grampositive antibiotics progressed significantly earlier than others. The multivariate analysis demonstrated that the use of anti-Gram-positive antibiotics was independently associated with reduced PFS and OS.226
More than 30% of CLLs can be classified based on their expression of stereotypic B-cell receptors (BCRs), strongly proposing that specific antigens are implicated in the onset of CLL. Unmutated CLLs, containing Ig heavy chain variable (IGHV) genes in germline configuration express low-affinity, poly- and self-reactive BCRs. Nevertheless, the antigenic specificity of CLLs with mutated IGHV-genes (M-CLL) is still elusive. In a study, Hogeeboom et al. reported a new subset of M-CLL, presenting stereotypic BCRs highly specific for β-(1,6)-glucan, a major antigenic determinant of yeasts and filamentous fungi. β-(1,6)-glucan binding depended on both the stereotypic Ig heavy and light chains, as well as on a definite amino acid in the IGHV-CDR3. Reversion of IGHV mutations to germline configuration decreased the affinity for β-(1,6)-glucan, suggesting that these BCRs are really affinity-selected for their cognate antigen. Moreover, CLL cells presenting these stereotypic receptors grow in response to β-(1,6)-glucan. With this data it is attracting to hypothesize on the possibilities for pathogen-targeted treatments for this group of subjects.227
CONCLUSION
The microbiome is currently accepted as a specific organ with separate metabolic abilities that surpass the liver’s metabolism by a factor of 100. The microbiome is able to influence hematologic malignancies via several ways, including directly through metabolites and toxins, or indirectly via the innate and adaptive immune system.228 However, a number of issues remain unresolved and only further research will clarify whether it is sufficient to administer a single species of bacteria to achieve results or whether it is better to give a mixture of microorganisms, or if by modifying an individual's microbial composition, we can improve the effectiveness of immunotherapy.229-230 In addition, FMT could help manage critical illness such as acute leukemias. In fact, in the critical care setting, several elements such as use of antibiotics, aberrant nutrition, bloodstream infections, bowel ischemia and abnormal bowel motility, strongly contribute to intestinal dysbiosis, and FMT therapy should be investigated.231,232 Further studies are needed to clarify the rationale of FMT for cancer management such as reconstruction of intestinal microbiota, amelioration of bile acid metabolism and modulation of immunotherapy efficacy.
Substances with probiotic and prebiotic capacities may represent a novel approach to change microbiota structure with beneficial effects on tumor development. Targeted treatment on the microbiome by pre-or probiotics may be used for tumor prevention and particular alterations of the microbiome may be implemented as an adjuvant treatment to augment the effectiveness of current tumor therapies of chemotherapy and immuno-therapy.
ACKNOWLEDGEMENTS
The authors would like to thank Dr. Clarissa Novello of the University of Aberdeen for the editing of the text.
DISCLOSURES
All authors declare no conflicts of interest. No funding or financial support was received.
REFERENCES